
Synthèse 5 : GRANDES ÉTAPES DU DÉVELOPPEMENT ANIMAL ET VÉGÉTAL

Ce cours en ligne synthétique reprend les points de programme suivants :
- Notion de développement embryonnaire et post-embryonnaire
- Grandes étapes du développement embryonnaire animal
- Grandes étapes du développement embryonnaire des végétaux
- Fiche méthodologique : notion de lignage et d’identité cellulaire
Synthèse 5 : GRANDES ÉTAPES DU DÉVELOPPEMENT ANIMAL ET VÉGÉTAL
Grandes étapes du développement animal et végétal
Cette introduction est consacrée à poser les bases du développement animal et végétal afin de pouvoir étudier les comportements et interactions cellulaires à l’œuvre dans l’organisme en développement (synthèses 6 à 9). L’enjeu (voir vos objectifs d’apprentissage sur cette synthèse) est que vous reteniez le vocabulaire de base préalable à la description de phénotypes in vivo et que vous soyez capables de situer quelques structures embryonnaires dans l’espace et le temps et de citer leur devenir.
Développement embryonnaire et post-embryonnaire
Il est assez étonnant de constater que quel que soit l'animal ou le végétal considéré, il a été initialement constitué d'une seule cellule, issue de la fusion entre un ovocyte et un spermatozoïde lors de la fécondation. Cette cellule unique initiale s'appelle le zygote. Le développement embryonnaire ou embryogenèse regroupe toutes les étapes qui permettent de transformer ce zygote en un organisme autonome avec une organisation tridimensionnelle précise correspondant à son plan d’organisation, c'est-à-dire avec des organes répartis correctement le long d'axes de polarité : par exemple l'axe antéro-postérieur, l'axe dorso-ventral et l'axe droite-gauche chez la plupart des animaux. Le développement des animaux se poursuit au cours de la période post-embryonnaire avec la croissance et la maturation des organes (notamment reproducteurs) et éventuellement une métamorphose, amenant un changement parfois radical de plan d'organisation (Figure 1). La croissance des animaux est généralement limitée dans le temps. Chez les végétaux elle est théoriquement illimitée. En outre l’organisme végétal se caractérise par une organogenèse continue grâce à l’activité des méristèmes que nous décrirons plus loin.
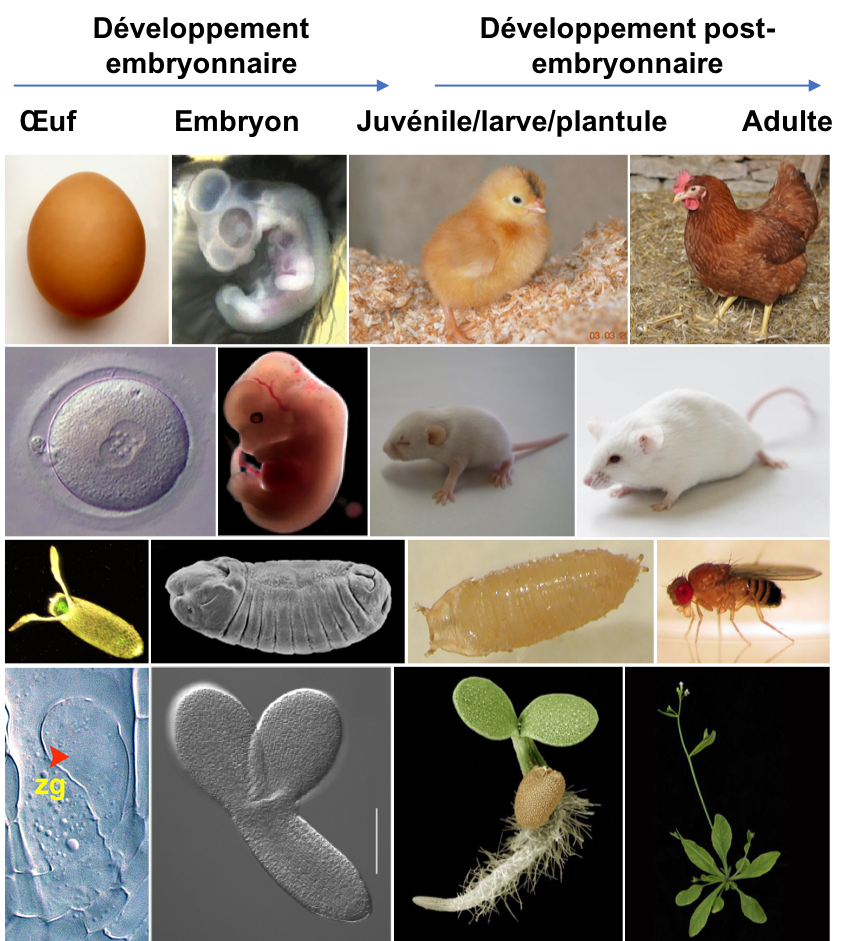
Figure 1 : Illustration du développement embryonnaire et post-embryonnaire chez 3 animaux (poulet, souris et drosophile) et un végétal (Arabidopsis). Le développement embryonnaire ou embryogenèse permet le passage de la cellule œuf à un individu (juvénile, larve, plantule) possédant les caractéristiques du plan d’organisation de l’espèce. Chez les animaux, il se poursuit par une période post-embryonnaire caractérisée par de la croissance, la maturation de certains organes (dont les organes reproducteurs) et une éventuelle métamorphose. C’est l’acquisition de la capacité à se reproduire qui caractérise l’état adulte. Chez les végétaux, croissance et organogenèse se poursuivent tout au long de la vie de l’individu.
Nous nous intéresserons ici essentiellement au développement embryonnaire. Il est constitué d'une succession d'étapes, largement conservées chez tous les animaux d'une part, et chez tous les végétaux d'autre part.
Le développement embryonnaire chez les animaux
Notion de feuillets embryonnaires
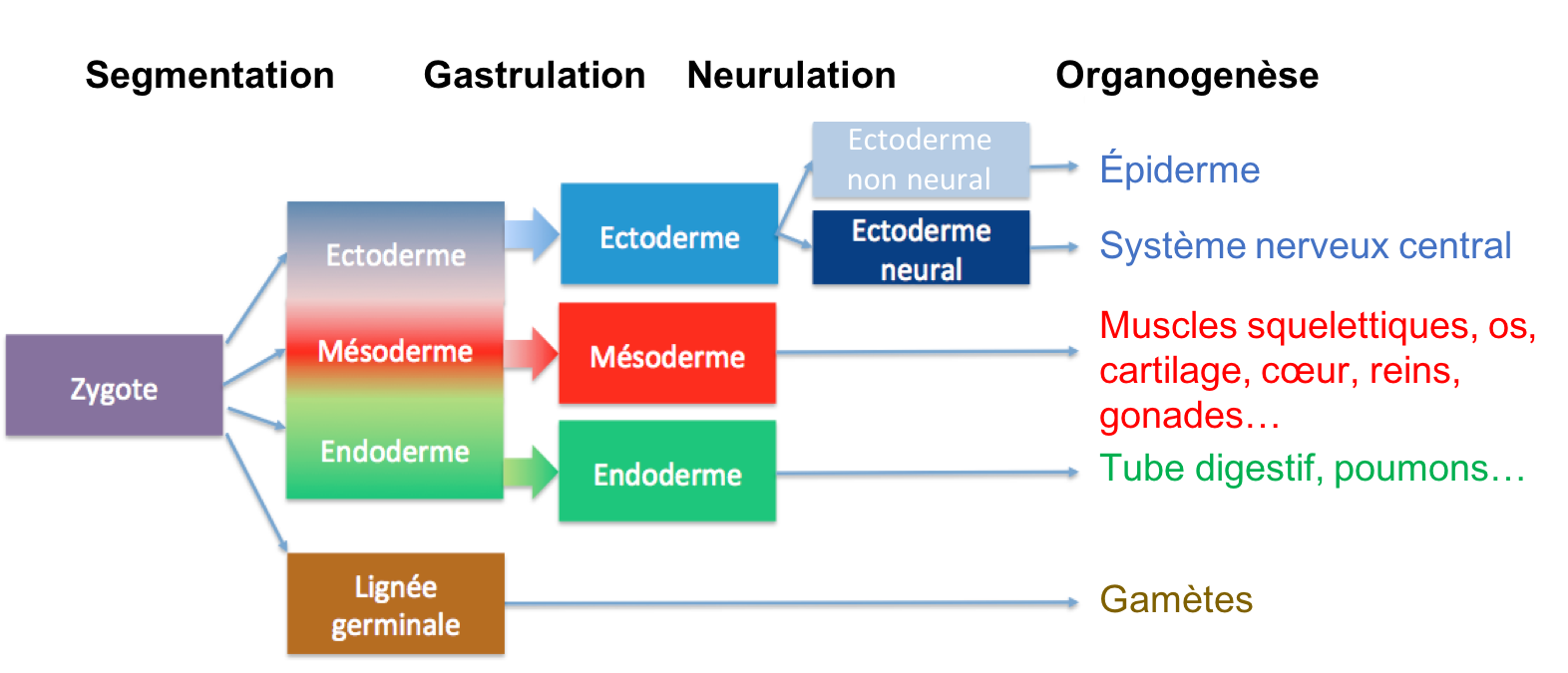
Pour passer du stade « zygote », cellule unique sphérique, à une structure tridimensionnelle pluricellulaire comprenant de multiples tissus et organes organisés dans l’espace selon le plan d’organisation de l’espèce, l’embryon va passer par différentes étapes faisant intervenir des divisions cellulaires, d’éventuels mouvements/migrations de cellules et de la différenciation cellulaire (voir synthèse 3). Tissus et organes ne se forment pas directement. Des groupes de cellules passent par des états intermédiaires et se différencient progressivement. Les premiers tissus (ou feuillets) embryonnaires sont au nombre de 3 chez les organismes bilatériens : ectoderme, mésoderme et endoderme. Ce sont les tissus fondateurs de l’ensemble des tissus adultes et chacun d’entre eux participe à la formation d'un ensemble de tissus et organes bien précis résumés dans le tableau ci-après (Tableau 1).
En plus des cellules des trois feuillets, notons la présence des cellules de la lignée germinale qui donnent naissance aux gamètes.
Les grandes étapes de l’embryogenèse animale
Les grandes étapes de l’embryogenèse animale sont les suivantes (Figure 2) :
- Clivage ou segmentation : première étape du développement embryonnaire où le zygote puis l'embryon subit des mitoses successives, ce qui divise le cytoplasme issu de l'ovocyte. L'embryon devient multicellulaire. C’est au cours du clivage que se forment les 3 feuillets embryonnaires (voir synthèse 7). Attention : les divisions cellulaires continuent dans les étapes ultérieures du développement, dans l’ensemble des tissus en formation. Elles contribuent aussi à la croissance de l’organisme au cours du développement post-embryonnaire.
- Gastrulation : phase du développement embryonnaire où d’importants mouvements cellulaires vont réorganiser les feuillets dans l’espace. À l'issue de la gastrulation, les axes de polarité de l’embryon commencent à être visibles. Attention : il existe des mouvements et migrations cellulaires dans les étapes ultérieures du développement, même s’ils ne sont pas aussi généralisés (voir synthèse 8).
- Neurulation : phase du développement embryonnaire (en fin de gastrulation ou juste après) où le système nerveux central s’individualise du reste de l’ectoderme.
- Organogénèse : phase du développement embryonnaire (incluant la neurulation mais se poursuivant à l’issue de cette dernière) au cours de laquelle les organes commencent à se former par interaction entre les différents tissus.
Exemple de l’embryogenèse du Xénope
Étudions maintenant ces
différentes étapes sur un organisme-modèle : le Xénope. Il s'agit d'un Amphibien Anoure (sans queue après la
métamorphose) qui est bien adapté à l'élevage. Après stimulation hormonale, une
femelle peut pondre plusieurs centaines d'ovocytes qui sont fécondés in vitro avec un broyat de testicules de
mâles. Les ovocytes et les embryons aux premiers stades font 2 mm de diamètre
et sont facilement observables à l’œil
nu ou à la loupe binoculaire. Le développement
embryonnaire est rapide (à 25°C) mais il peut être ralenti si besoin en
incubant les embryons à des températures plus basses (18°C voir 12°C).
Commencez par regarder ce film où vous allez retrouver la succession des étapes
développementales décrites ci-dessus :
La figure 3 résume l’ensemble des étapes observées :
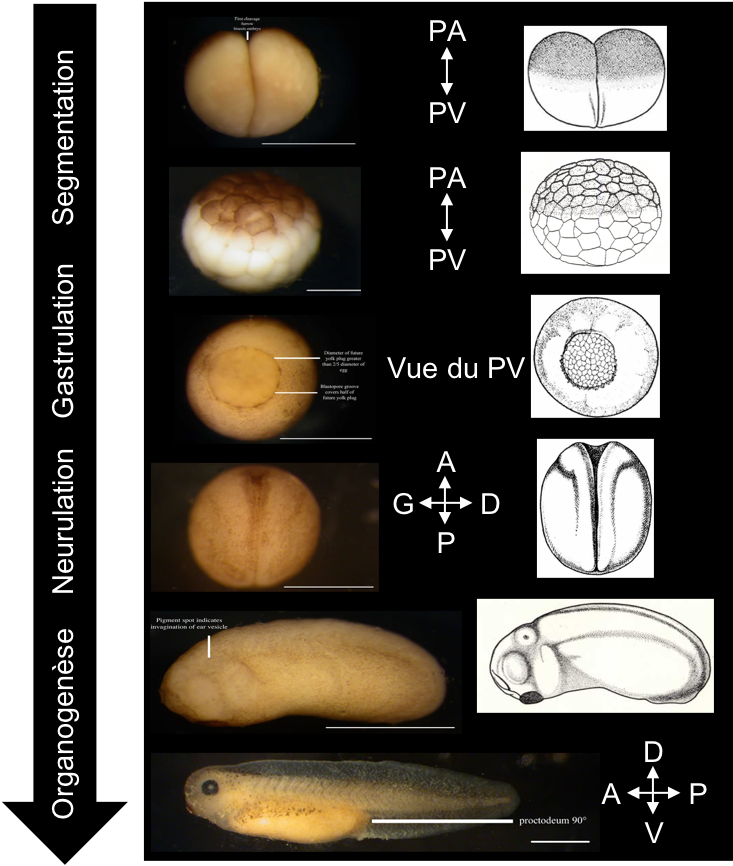
Figure 3 : Étapes du développement embryonnaire chez le Xénope. À cette échelle, vous observez :
- Des divisions cellulaires sans augmentation de la taille de l’embryon (clivage ou segmentation).
- L’apparition d’une fente et des mouvements tissulaires (gastrulation).
- Le soulèvement puis la fermeture de tissus dorsaux (neurulation).
- L’allongement de l’embryon dans l’axe antéro-postérieur et l’apparition d’organes (dont yeux, plaque adhésive, branchies; organogenèse): le plan d’organisation devient visible.
Allez maintenant sur la page
suivante : https://view.genial.ly/5c0ce7510fd38247366d9537/developpement-xenope-l2
Une image interactive retraçant le cycle de vie du Xénope vous permet de suivre étape par étape son développement embryonnaire. Voici les points qui doivent particulièrement retenir votre attention :
- Observez la polarisation de l'ovocyte et ses réserves indispensables au développement embryonnaire (vitellus).
- Notez les mouvements du cytoplasme cortical qui adviennent au moment de la fécondation (rotation corticale et formation du croissant gris). Nous reviendrons sur ce phénomène et son importance en voir synthèse 7.
- Notez la particularité des premières divisions de la segmentation (cycles dépourvus de phases G1 et G2) et le fait que le génome du zygote ne commence à s’exprimer que lorsque les cycles redeviennent classiques (au moment de la transition mid-blastuléenne). Tous les ARN et protéines nécessaires au début du développement ont été hérités par la femelle via l’ovocyte. Ils ont été synthétisés au cours de l’ovogenèse.
- Retenez le principe de l’établissement d’une carte des territoires présomptifs.
- Observez les mouvements cellulaires qui ont lieu au cours de la gastrulation et leurs conséquences sur la réorganisation spatiale des trois feuillets (le détail de ces mouvements n’est pas à connaître).
- Observez comment le tube neural se forme.
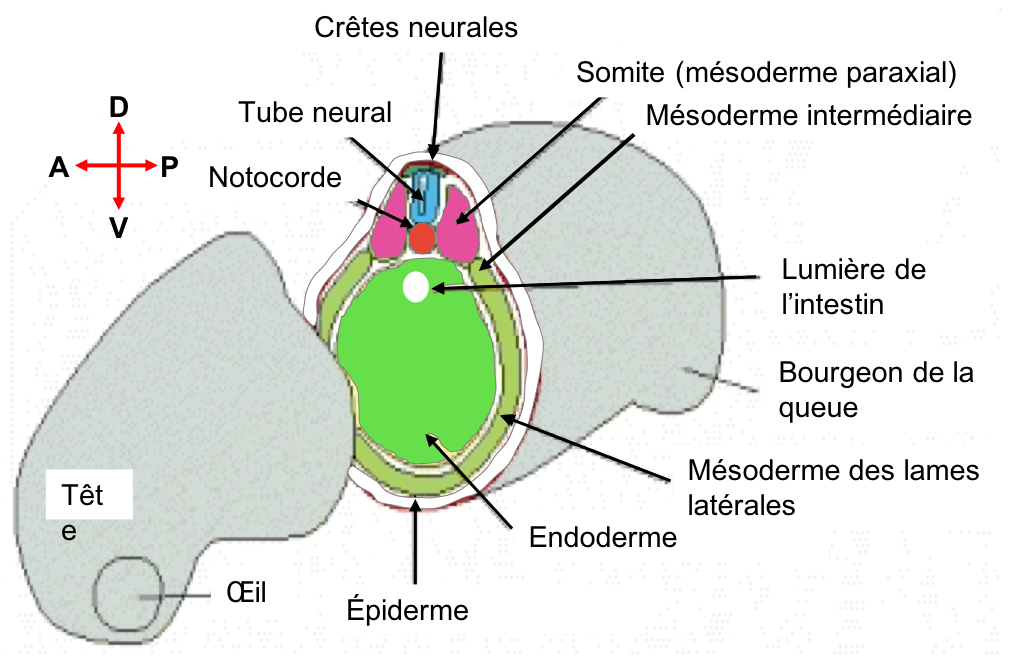
- Familiarisez-vous avec une coupe d’embryon au stade bourgeon caudal (voir également Figure 4).
- Observez la formation des somites à partir du mésoderme paraxial et leur régionalisation. Nous reviendrons sur ce processus en synthèse 7.
Pour
aller plus loin et découvrir de façon animée et commentée le développement embryonnaire
du Xénope, regardez la vidéo suivante :
Les étapes du développement embryonnaire chez les végétaux
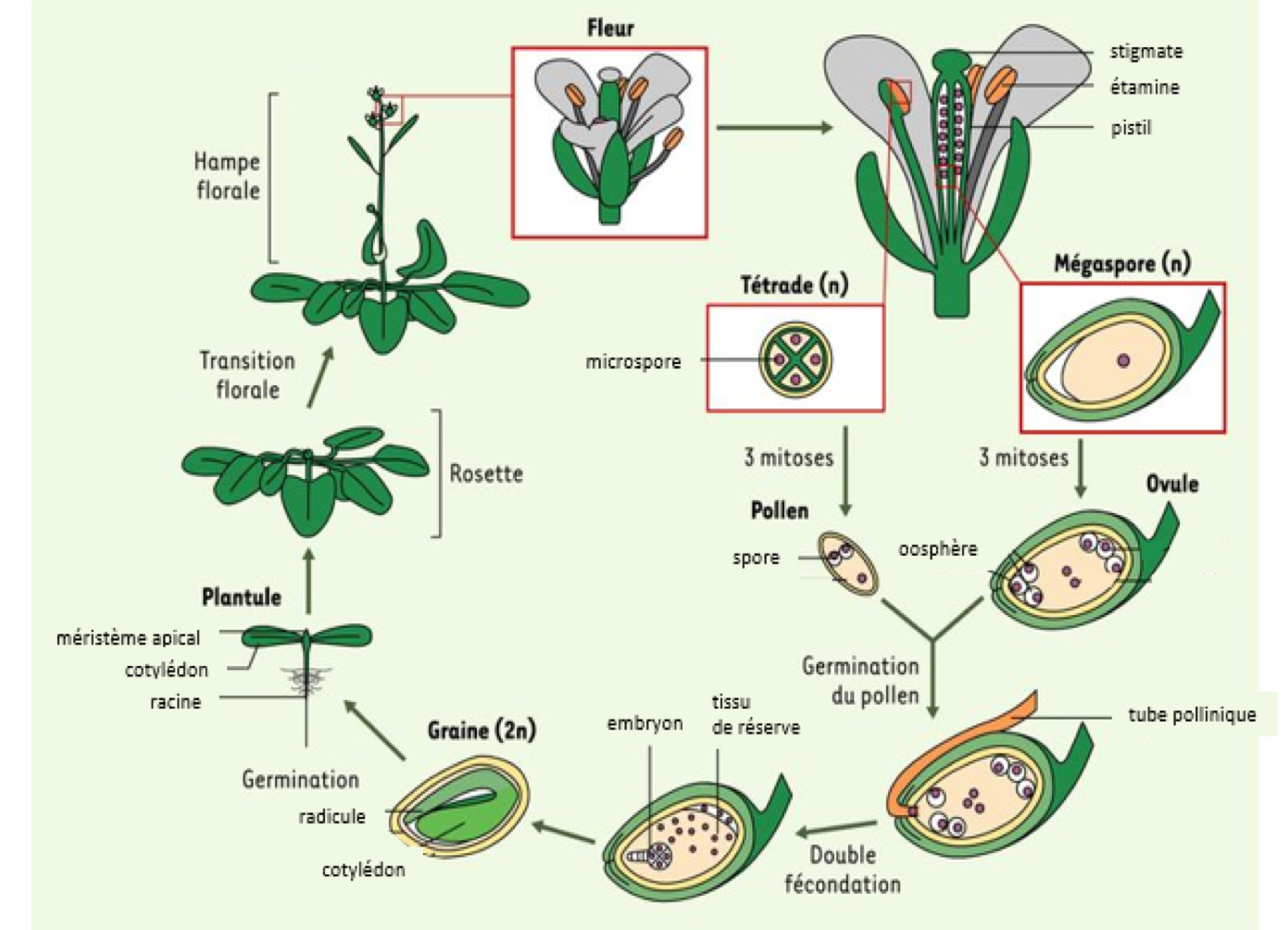
Le
développement embryonnaire démarre avec la fécondation d'une oosphère (gamète
femelle) par un gamète mâle qui est issu du grain de pollen. L'embryon végétal
passe ensuite d'un stade à un autre, nommés d'après leur nombre de cellules ou
la forme générale de l'embryon. Du zygote jusqu'au stade embryon mature, le
développement prend environ deux semaines. Observez ces deux petits
films. Le premier est un film accéléré du développement embryonnaire d’Arabidopsis
et le second vous montre la dissection d’un embryon à partir de graines
extraites d’une silique (activer les sous-titres) :
Observez maintenant la Figure 6. On distingue 2 grandes structures dans l’embryon :
- Un tissu extra-embryonnaire dans la partie basale de l’embryon, nommé suspenseur. Ce dernier relie embryon et tissus maternels, assurant un approvisionnement en ressources nutritives. Sa prolifération pousse l'embryon vers le centre de la graine.
- L’embryon stricto sensu dans la partie apicale, appelé proembryon. C’est lui qui est à l’origine de la quasi-totalité des différents tissus de la plantule.
Ces deux structures commencent à
se former dès la première division du zygote. Cette division est asymétrique et
produit une petite cellule, la cellule apicale,
et une grande cellule, la cellule basale. C'est
une division fondamentale car tout l'axe apico-basal
de la plante en découle. La cellule basale va poursuivre ses divisions pour
donner le suspenseur et une petite partie du
proembryon appelée hypophyse. Tout le reste du
proembryon est issu de la cellule apicale.
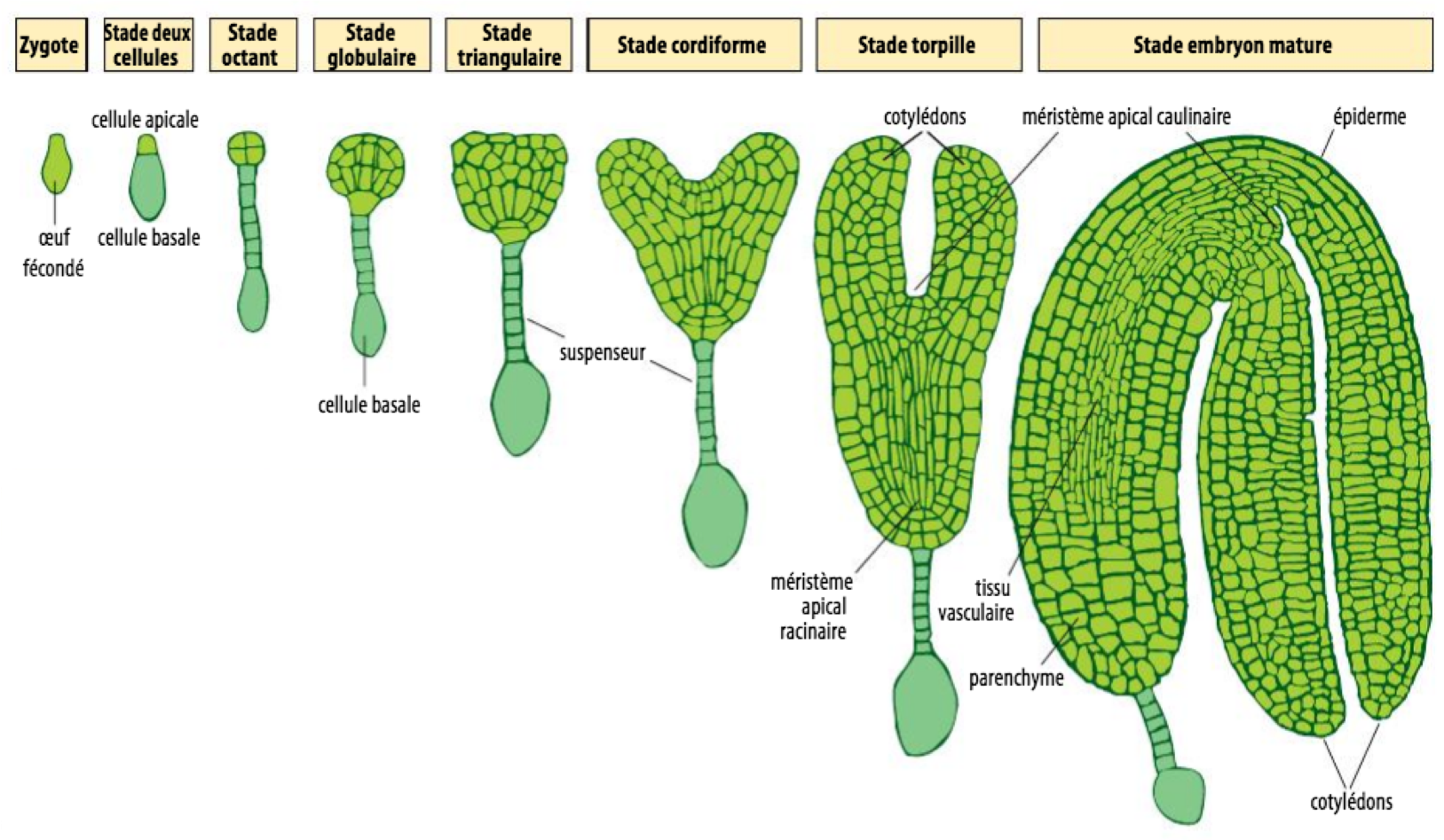
Figure 6 : Stades du développement embryonnaire d'Arabidopsis thaliana. La première division du zygote est asymétrique et génère une petite cellule apicale et une grande cellule basale. Cette dernière participe à la formation du suspenseur (structure extra-embryonnaire qui a une fonction nutritive) et aussi à l'hypophyse qui générera une partie du méristème racinaire de l’embryon. La cellule apicale donne naissance au reste de l'embryon et se divise jusqu'à former une masse globulaire qui donne son nom au stade correspondant. L'embryon commence à s'allonger selon l'axe apico-basal tandis qu'émergent les cotylédons (premières feuilles). C'est le stade cordiforme (en forme de cœur) puis le stade torpille. L'embryon mature est ensuite replié dans la graine. Il est en vie ralentie et reprendra son développement à la germination pour former la plantule. Source : Wolpert, Biologie du Développement : les grands principes. Dunod 2004.
Observez maintenant les différentes zones du proembryon et leur devenir (Figure 7). Au stade cœur (ou embryon cordiforme), des protubérances latérales apparaissent formant les deux pointes supérieures du cœur : ce sont les cotylédons, premières feuilles de la plantule. La présence de deux cotylédons permet de classer Arabidopsis parmi les Dicotylédones (les Monocotylédones tels que le maïs ou le blé n’en possèdent qu'un seul). L'hypocotyle, jeune tige, et la racine se mettent ensuite en place.
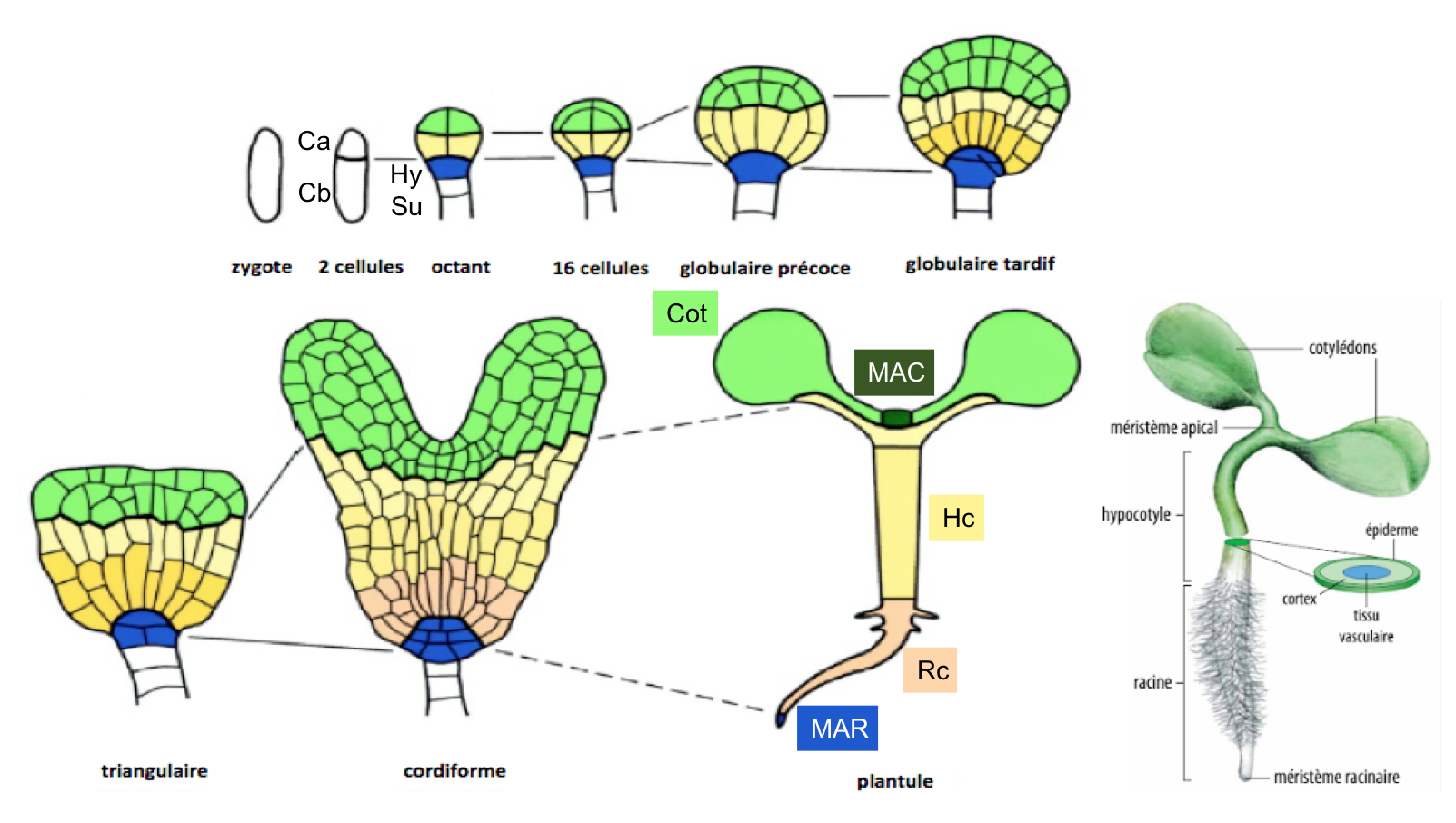
Figure 7 : Devenirs des différents territoires du proembryon d'Arabidopsis thaliana. Des divisions cellulaires très stéréotypées donnent naissance à divers territoires embryonnaires, représentés ici par un code couleur. La cellule apicale génère l'ensemble des structures de la plantule à l’exception d’une partie du méristème racinaire qui dérive de la cellule basale. Ca : cellule apicale; Cb : cellule basale; Hy : hypophyse; Su : suspenseur; Cot : cotylédon; MAC : méristème apical caulinaire; Hc : hypocotyle; Rc : racine; MAR : méristème apical racinaire. Référence : Laux et al. (2004) Plant Cell, 16, 190-202.
L’embryogenèse
a également mis en place deux structures fondamentales pour la suite du
développement : le méristème apical caulinaire (MAC)
et le méristème apical racinaire (MAR). Un
méristème est un tissu constitué de cellules indifférenciées formant une zone de croissance où des divisions cellulaires vont se
poursuivre tout au long de la vie de la plante. En générant de nouvelles
cellules qui se différencieront, le MAC et le MAR
assurent ainsi la croissance en longueur de la partie aérienne de la plante et
de sa racine respectivement (Figure 8). Le
MAR est uniquement histogène (il ne génère que de nouvelles cellules dans la
racine) tandis que le MAC est histogène (genèse de cellules de la tige) et
organogène (genèse de nouvelles feuilles et bourgeons et de fleurs après la
transition florale). Ce sont des méristèmes dits « primaires » dont nous
reverrons plus précisément la structure et le fonctionnement en en synthèse 9.
Chez les plantes ligneuses, des méristèmes dits « secondaires »
assurent la croissance en épaisseur de la plante.
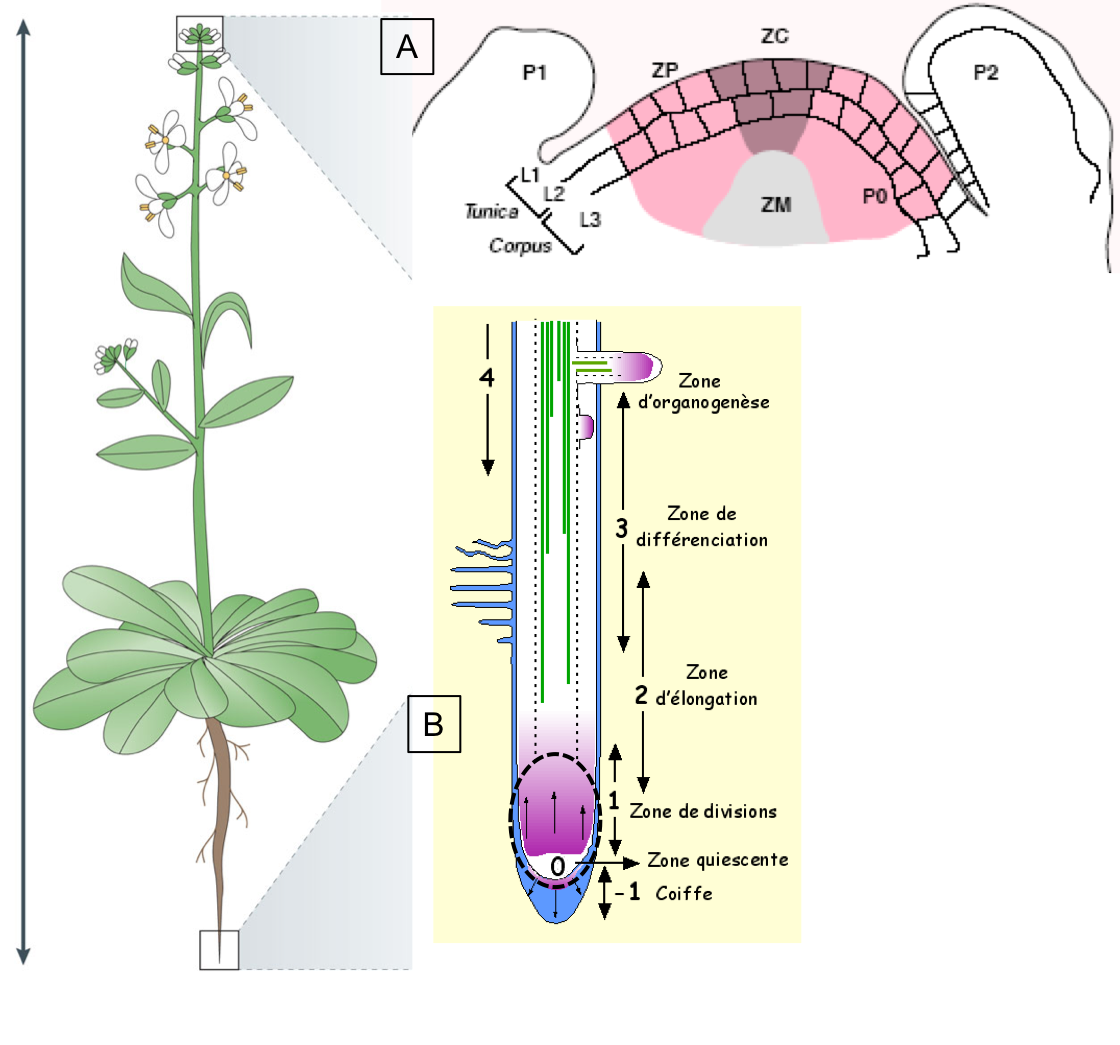
Figure 8 : Structure des méristèmes apicaux caulinaire (A ; MAC) et racinaire
(B ; MAR). (A) Schéma d’une
coupe effectuée au niveau du MAC. Le MAC contient plusieurs couches de cellules
notées L1, L2 et L3. L1 et L2 forment la Tunica ; L3 est appelée Corpus. Ces
couches sont définies par les plans de divisions des cellules qui les
composent : divisions anticlines dans la tunica
et divisions sans plan défini dans le Corpus. On peut également distinguer des zones sur
la base de l’intensité de prolifération cellulaire observée : divisions
lentes et peu fréquentes dans la zone centrale (ZC) ; divisions rapides et
fréquentes dans la zones périphérique (ZP ; lieu d’initiation des organes
latéraux) ; vitesse intermédiaire de division dans la zone médullaire
(ZM ; zone de formation des tissus internes de la tige). P = primordium de
jeune feuille nouvellement formée. (B)
Schéma d’une coupe de racine. Le MAR est entouré en noir. Il comprend un centre
quiescent (0 ; cellules qui se divisent peu) et une zone de division (1). Les
cellules qui sont générées par le MAR vont ensuite croître (2 ; zone
d’élongation) et se différencier (3 ; zone de différenciation). Dans la
zone 4 dite « d’organogenèse » des racines latérales peuvent se
former.