
Synthèse 1 : STRUCTURES ET COMPOSANTS CELLULAIRES
 Ce cours en ligne synthétique reprend les points de programme suivants :
Ce cours en ligne synthétique reprend les points de programme suivants :
- Synthèse sur le cytosquelette
- Synthèse sur la matrice extracellulaire animale
- Synthèse sur la paroi végétale
Introduction à la matrice extracellulaire animale
Dans les tissus des organismes pluricellulaires animaux, l’espace extracellulaire est occupé par la matrice extracellulaire (MEC). Cette dernière est constituée à 70% d’eau et d’un réseau complexe et très organisé de macromolécules, sécrétées par les cellules environnantes (Figure 1). Toutes les cellules eucaryotes des métazoaires produisent une MEC. Pour certaines cellules, comme les fibroblastes des tissus conjonctifs ou les cellules osseuses et cartilagineuses, il s’agit là de leur fonction principale. À chaque type cellulaire est associée une MEC spécifique dans sa composition et son organisation. La structure des MEC peut en outre évoluer en fonction de différentes situations, physiologiques (développement/croissance ou remodelage du tissu au cours de la cicatrisation), ou pathologiques (fibroses, ostéoarthrose ...).
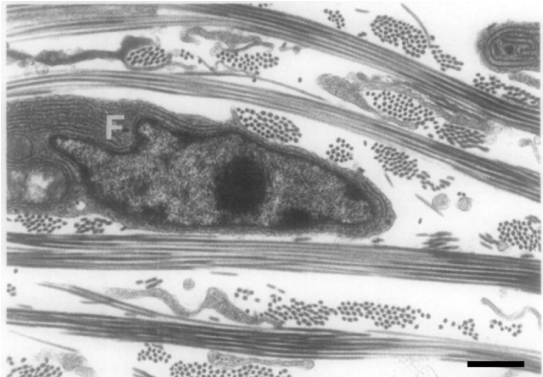
Figure 1 : Fibroblaste entouré de sa matrice dans une coupe de derme d’embryon de poulet. Photographie prise au microscope électronique à transmission. Les fibrilles de collagènes sont ordonnées en faisceaux organisés de façon orthogonale, parallèlement à la surface du derme. Embryon de 15 jours. Barre d’échelle : 1 micron. D’après Ploetz et al, 1991 Structural Biology 106, 73-81.
Fonctions des matrices extracellulaires
Les MEC jouent un rôle essentiel dans l’architecture tissulaire en constituant un ciment entre les cellules et en assurant la résistance mécanique des tissus. Outre cette fonction structurale, nous aurons l’occasion de voir au cours de l'UE qu’elles sont également des acteurs majeurs de la régulation des grandes fonctions cellulaires comme la prolifération, la différenciation ou la migration. Nous verrons par quels mécanismes en nous penchant sur leur rôle dans la signalisation cellulaire (synthèse 2).
Structure des matrices extracellulaires
La grande diversité des matrices extracellulaires est liée au grand nombre de molécules qui la composent, et à leurs multiples capacités d’interaction.
La matrice extracellulaire est composée d'un ensemble de polysaccharides et de protéines. Les polysaccharides sont des glycosaminoglycanes (GAG) qui sont généralement reliés de façon covalente à une protéine pour former des « protéoglycanes ». Dans la composante protéique de la matrice, on distingue : un premier groupe, constitué par le collagène et l'élastine, essentiellement responsables de la structure de la matrice et un second groupe, moins abondant, constitué par la fibronectine et la laminine. Ce dernier est plutôt impliqué dans l'organisation de sa structure (formation d'une trame) et dans l'adhérence cellule-matrice.
Les chaines de glycosaminoglycanes
- Les glycosaminoglycanes sont de longues chaînes composées d'unités disaccharidiques répétitives. Un des deux résidus glucidiques est toujours un glucide aminé (N-acétylglucosamine ou N-acétylgalactosamine) et le second est habituellement un acide uronique (glucuronique ou iduronique). Les GAG se répartissent en quatre types principaux :
o acide hyaluronique
o chondroïtine sulfate
o héparane sulfate
o kératane sulfate
- À l'exception de l'acide hyaluronique, tous les GAG sont liés de façon covalente à une protéine pour former des protéoglycanes (Figure 2).
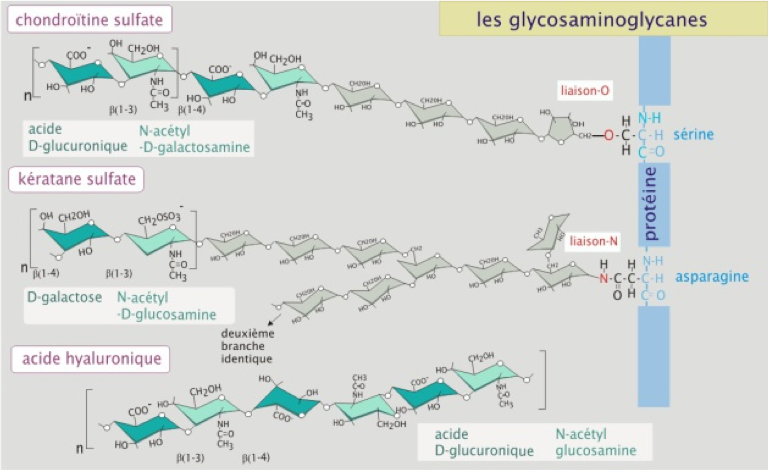
Figure 2 : Structure des GAG.
- Les GAG, et particulièrement l'acide hyaluronique, forment des mailles qui permettent de retenir de grandes quantités d'eau. Ils constituent un gel d'un volume considérable (1 000-10 000 fois le volume initial) qui permet à la matrice de résister aux forces de compression.
- Les GAG ont aussi un rôle essentiel dans la migration cellulaire au cours de la morphogenèse et de la cicatrisation des tissus. En cas de blessure, après la coagulation du sang, le premier produit qui entre dans la lésion est l'acide hyaluronique. Il forme un échafaudage qui va servir de support aux cellules leucocytaires et aux fibroblastes pour reconstruire le nouveau tissu.
Le collagène et l’élastine
- Le collagène représente environ un quart de la masse protéique d'un organisme humain. Le collagène est la protéine structurale majeure des MEC qui forme des armatures, renforçant les structures telles que les tendons, la peau et les organes internes.
- Les collagènes forment une famille multigénique de protéines fibreuses. Tous les collagènes sont constitués de trois protéines (appelées chaînes alpha) associées les unes aux autres par des liaisons hydrogène. Les trois chaînes alpha peuvent être identiques (homotrimériques) ou différentes (hétérotrimériques). Elles s’organisent en s’enroulant les unes autour des autres pour former une triple hélice (la molécule de tropocollagène). Les triples hélices s’associent pour former des structures d’ordre supérieur, les fibrilles de collagène (environ 0,5 microns de diamètre) puis les fibres (plusieurs microns de diamètres ; Figure 3).Contrairement aux GAG, qui résistent aux forces de compression, les fibres de collagène forment des structures qui résistent aux forces de tension.
- L’élasticité de certains tissus est quant à elle assurée par un réseau de molécules matricielles fibreuses incluant l’élastine.
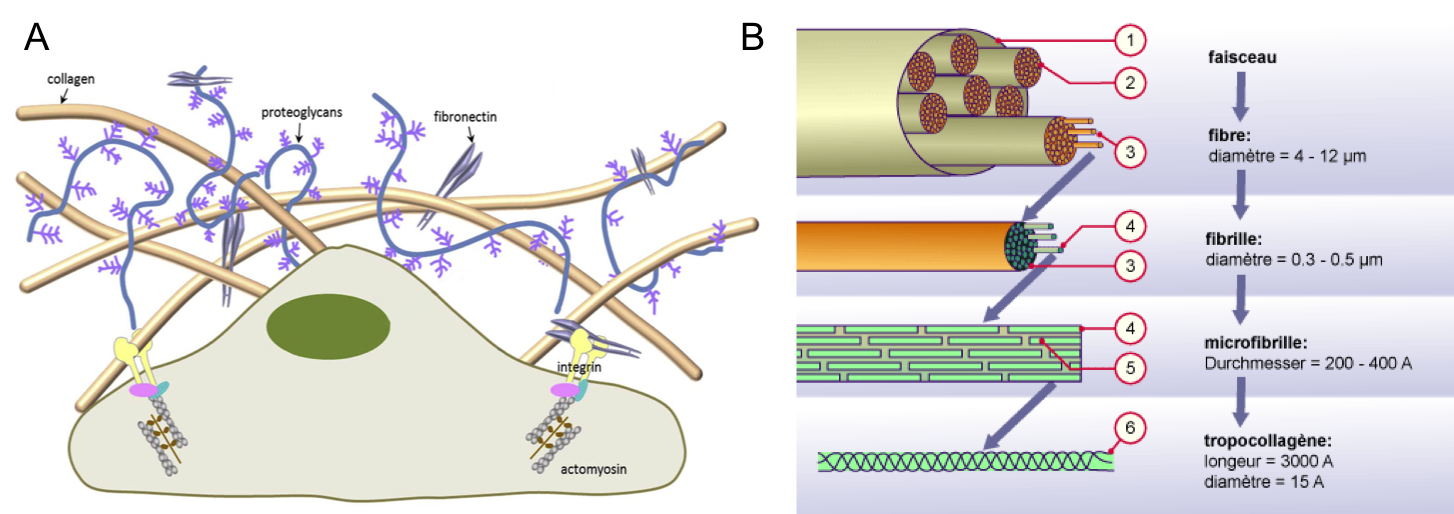
Figure 3 : (A) Représentation schématique des composants de la matrice extracellulaire. D’après Thannickal et al, 2014. https://ajp.amjpathol.org/article/S0002-9440(14)00156-4/fulltext. (B) Assemblage du tropocollagène en structures d’ordre supérieur (1. Faisceau ; 2. Fibre ; 3. Fibrille ; 4 : Microfibrille ; 5 : Tropocollagène ; 6 : Chaîne alpha).
La fibronectine
- La fibronectine forme un réseau fibrillaire qui contribue à organiser la matrice et à favoriser l'adhérence des cellules à cette dernière.
- Une molécule de fibronectine est constituée de deux sous-unités α et β réunies par une paire de ponts disulfure. Il existe une vingtaine d’isoformes de fibronectine, produites par épissage alternatif du même gène.
- La fibronectine contient différents domaines qui constituent des sites d'interactions spécifiques avec d'une part les constituants moléculaires de la matrice extracellulaire (formation d'une trame) et d'autre part les cellules (par l'intermédiaire de récepteurs nommés « intégrines » ; voir synthèse 2).
- La fibronectine joue un rôle important dans le guidage des cellules lors des migrations embryonnaires chez les vertébrés (gastrulation et migration des cellules de la crête neurale).
La laminine
- La laminine est un gros complexe protéique flexible constitué de trois chaînes polypeptidiques.
- Douze formes différentes de chaînes de laminine ont été caractérisées.
- À l’instar de la fibronectine, la laminine peut interagir avec différents composants matriciels et avec les cellules elles-mêmes.
- La laminine est le constituant essentiel des lames basales, MEC sur laquelle reposent tous les feuillets épithéliaux et endothéliaux (entoure également les cellules musculaires et adipeuses) et qui permet notamment leur adhérence aux tissus conjonctifs sous-jacents.